A Deep Dive into GLP-1 Tissue Expression
Disclaimer: The information provided in this article is for educational and informational purposes only. It is not intended as medical advice, diagnosis, or treatment. Products and compounds referenced are for research use only and are not approved for human or veterinary consumption. Always consult a qualified professional regarding health or medical decisions.
Why Understanding GLP-1 Receptor Expression Tissues Matters for Modern Research

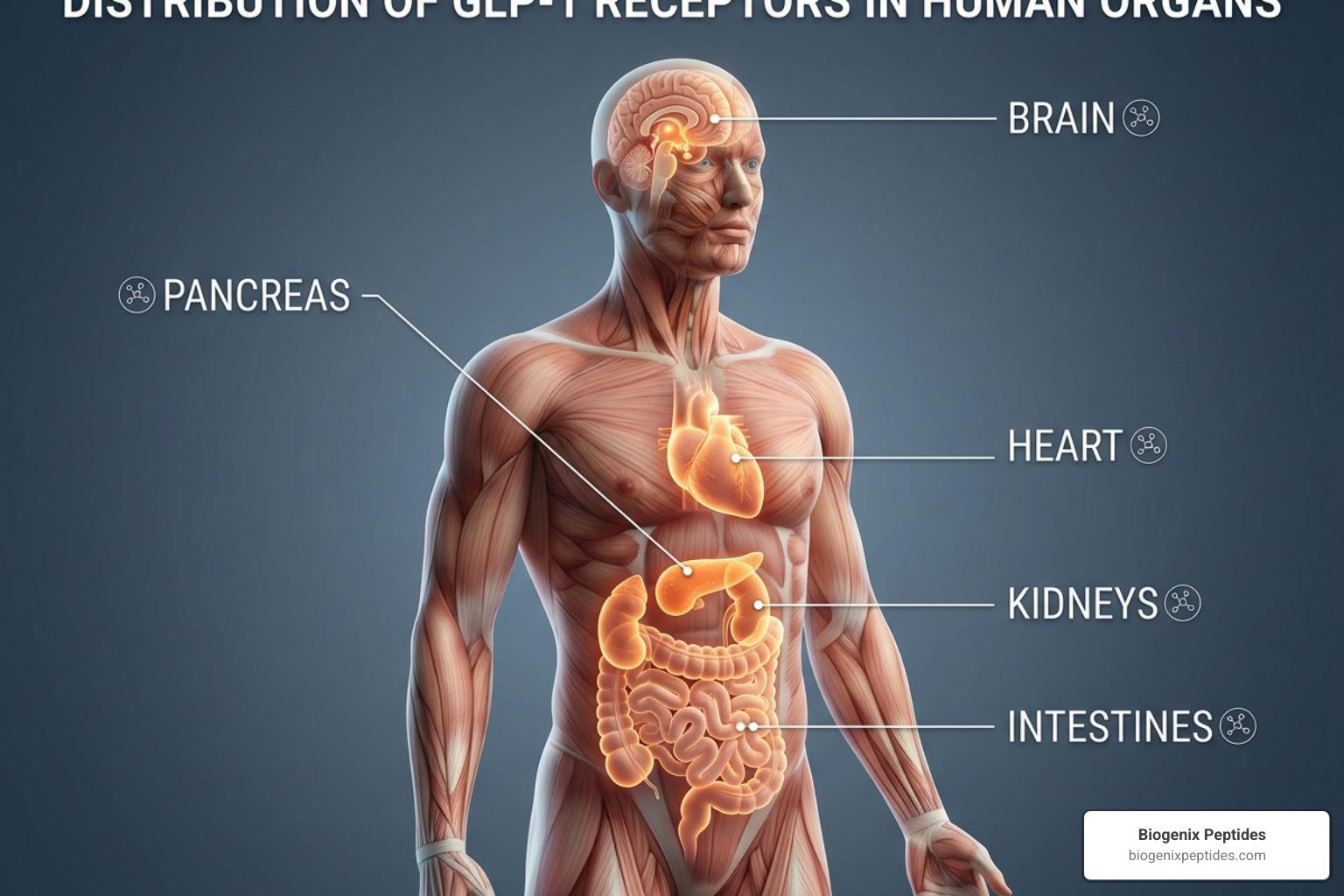
GLP-1 receptor expression across tissues is far more widespread than most people expect. This receptor isn’t just a pancreatic switch — it shows up in the brain, heart, kidneys, gut, and even certain tumors.
Here’s a quick snapshot of where GLP-1R is found and at what level:
| Tissue / Cell Type | Expression Level | Notes |
|---|---|---|
| Pancreatic beta cells | High | Primary site; drives insulin secretion |
| Tuft cells (GI tract) | Very high (46.1 nCPM) | Highest single-cell expression recorded |
| Supraoptic nucleus (brain) | Notable (8.2 nTPM) | Highest brain region expression |
| Heart (sinoatrial node) | Moderate | Myocytes only; not ventricular cardiomyocytes |
| Brunner’s glands (duodenum) | High | Strongest GI mucosal expression |
| Kidney / Lung | Low-moderate | Restricted to arterial smooth muscle cells |
| Liver, skeletal muscle, adipose | Absent | Not detected in these tissues |
| Pheochromocytomas (tumor) | Very high | Highest expression among all tumors tested |
GLP-1R tissue specificity Tau score: 0.91 — indicating strong tissue-enriched expression rather than ubiquitous distribution.
Understanding this distribution matters because GLP-1 receptor agonists are among the most widely used research compounds in metabolic science today — with global drug sales reaching $22 billion in 2022. Yet the exact tissues and cell types that respond to GLP-1R activation are still being mapped with precision. Early studies were hampered by non-specific antibodies, leading to conflicting reports. More recent work using validated monoclonal antibodies, RNAscope mRNA detection, and SNAP-tag transgenic models has started to give researchers a clearer picture.
This guide breaks down the current evidence — tissue by tissue, cell type by cell type — using data from receptor autoradiography, RNA-seq, single-cell transcriptomics, and immunohistochemistry.
I’m Jay Daniel, Founder and CEO of BioGenix Peptides, and my work in peptide research has given me a deep interest in how receptor-level biology — including GLP-1 receptor expression across tissues — shapes the way we understand and develop peptide-based research tools. In the sections ahead, we’ll walk through each major tissue system in detail, so you can follow the science wherever it leads.

Simple guide to GLP-1 receptor expression tissues terms:
- GLP-1 receptor agonist benefits
- GLP-1 agonists cardiovascular outcomes
- GLP-1 agonists muscle preservation
Mapping GLP-1 Receptor Expression Tissues: A System-Wide Overview
When we look at GLP-1 receptor expression tissues on a macro scale, we find a highly specific, tissue-enhanced profile. According to transcriptomic databases, the GLP-1 receptor (GLP1R) is classified as a G-protein coupled receptor (GPCR) and is a primary target of clinical interest. It has a high Tau specificity score of approximately 0.90 to 0.91, indicating that it is not uniformly distributed throughout the body but is concentrated in specific anatomical niches.

At the protein level, robust evidence confirms that the receptor is primarily expressed in the pancreas, specific regions of the central nervous system, and key segments of the gastrointestinal tract. It is also found in lower but functionally significant amounts in the kidneys, lungs, and heart. Conversely, extensive profiling has shown that the GLP-1 receptor is completely absent in the liver, skeletal muscle, and adipose tissue. This absence is a crucial detail for researchers studying metabolic pathways, as it indicates that GLP-1’s profound effects on hepatic glucose production and muscle insulin sensitivity are mediated indirectly through systemic signaling rather than direct receptor binding in those specific organs.
To explore the detailed transcriptomic data across various organs, researchers can refer to the Tissue expression of GLP1R – Summary – The Human Protein Atlas. Understanding these system-wide dynamics helps clarify Why Your Doctor Is Obsessed with GLP-1 Receptor Agonists and why their physiological impact extends far beyond simple glycemic control.
Cellular Heterogeneity of GLP-1 Receptor Expression Tissues in the Pancreas
The pancreas is the historical home of GLP-1 research. It is here that GLP-1 plays its classic role in the incretin effect, which, alongside GIP, accounts for up to 70% of meal-stimulated insulin secretion. However, looking at the pancreas as a single, uniform tissue misses the incredible cellular diversity within it.
Within the pancreatic islets of Langerhans, GLP1R expression is highly heterogeneous:
- Beta Cells: This is where the densest concentration of GLP-1 receptors resides. Activation of these receptors on beta cells stimulates adenylyl cyclase, raising intracellular cyclic AMP (cAMP) and triggering glucose-dependent insulin secretion.
- Delta Cells: Recent transgenic reporter models have demonstrated that delta cells (which secrete somatostatin) also express functional GLP-1 receptors. This suggests a local, paracrine feedback loop where GLP-1 directly influences somatostatin release to modulate nearby beta and alpha cell activity.
- Alpha Cells: Interestingly, only a tiny fraction of glucagon-secreting alpha cells show GLP1R expression. Studies in rodent models indicate that only about 8.3% of alpha cells exhibit faint GLP1R overlap, suggesting that GLP-1’s ability to suppress glucagon secretion is largely an indirect process mediated by somatostatin and insulin.
- Acinar Cells: Outside of the endocrine islets, weak but detectable GLP-1R expression is observed in the exocrine acinar tissue. Crucially, pancreatic ductal epithelial cells show no GLP-1R expression, resolving a long-standing debate regarding whether GLP-1 therapies directly stimulate ductal proliferation.
Advanced imaging techniques have allowed us to peer deeper into this cellular landscape. Researchers using genome-edited models have discovered that endogenous GLP-1R expression and signaling are highly dynamic. For instance, studies published in Revealing the tissue-level complexity of endogenous glucagon-like peptide-1 receptor expression and signaling | Nature Communications show that different receptor agonists recruit distinct subpopulations of islet cells into signaling and internalization pathways, highlighting a complex layer of cellular cooperative behavior.
Central Nervous System and Vagal Ganglia Distribution
The profound effects of GLP-1 receptor agonists on appetite suppression, satiety, and food reward are directed by the brain. The GLP-1 receptor is expressed in highly specific regions of the central nervous system (CNS) and the peripheral nervous system.
In the brain, the highest density of GLP1R is found in the hypothalamus and the brainstem. Specifically, the supraoptic nucleus shows the highest regional expression at approximately 8.2 nTPM. Other key sites include:
- The Arcuate Nucleus (ARC): A critical hub for energy homeostasis where GLP-1R activation directly influences pro-opiomelanocortin (POMC) neurons to promote satiety.
- The Paraventricular Nucleus (PVN): Involved in coordinating metabolic and autonomic responses.
- The Area Postrema (AP) and Nucleus of the Solitary Tract (NTS): Located in the hindbrain, these regions lie outside the tightest parts of the blood-brain barrier, making them highly accessible to circulating GLP-1 and its therapeutic mimetics.
- The Neurohypophysis: Interestingly, the posterior pituitary (neurohypophysis) exhibits the absolute highest GLP-1 receptor density among all normal human tissues measured in autoradiography studies.
Beyond the brain itself, the vagus nerve serves as a vital communication highway between the gut and the brain. The nodose ganglion, which houses the cell bodies of vagal sensory neurons, is a major site of GLP1R expression. These vagal sensory neurons innervate the gastrointestinal tract and respond to both mechanical distension and locally released GLP-1.
A fascinating species difference has been uncovered here: in mice, Glp1r expression in the nodose ganglion is strictly neuronal. However, in human tissue, as detailed in Glucagon‐Like Peptide‐1 Targets in the Human Nodose Ganglion – PMC, Glp1r mRNA is expressed not only in about 7% of neurons at moderate-to-high levels (and 28% at low levels) but also in nonneuronal cells within the perineurium, epineurium, and connective tissue fascicles of the vagus nerve. This nonneuronal expression in humans may have unique implications for how human nerves respond to therapeutic agents.
Cardiovascular and Renal Expression Profiles
The cardiovascular benefits of GLP-1 receptor agonists have transformed them from simple metabolic tools into cornerstone therapies for heart health. To understand these systemic effects, we must look at where these receptors are located within the cardiovascular and renal systems.
In the heart, GLP-1R is localized to the myocytes of the sinoatrial node (the heart’s natural pacemaker). This specific localization explains why clinical trials and animal models consistently show a modest increase in resting heart rate upon receptor activation. Crucially, extensively validated monoclonal antibody mapping has shown that GLP-1 receptors are not present on ventricular cardiomyocytes. This indicates that the cardioprotective and remodeling benefits observed in cardiovascular studies are likely mediated through indirect pathways, such as improved endothelial function, reduced systemic inflammation, or altered sympathetic nervous system activity.
In the vascular system, GLP-1R expression is highly restricted. It is found exclusively in the smooth muscle cells of arteries and arterioles within the kidneys and the lungs. In the kidneys, this localization to renal arterial smooth muscle cells suggests a role in modulating renal blood flow and glomerular filtration rates, which may contribute to the observed nephroprotective effects of these compounds.
Researchers interested in the broader cardiovascular implications of these pathways can read more about how these mechanisms translate to clinical settings in Why GLP-1 Agonists Are a Change of Heart for Cardiac Care.
Subcellular Localization and Molecular Signaling Dynamics
At the microscopic level, the GLP-1 receptor is a dynamic protein that does not simply sit static on the cell surface. It is primarily localized to the plasma membrane, where it is poised to bind circulating ligands, but it is also found in intracellular compartments including nuclear bodies, the Golgi apparatus, and the centrosome.

As a Class B G-protein coupled receptor, the primary transduction pathway of GLP-1R is coupling to the Gs family of G-proteins. When a ligand binds to the extracellular domain:
- The receptor undergoes a conformational change that activates intracellular Gs proteins.
- This activation stimulates adenylyl cyclase, leading to a rapid conversion of ATP into cyclic adenosine monophosphate (cAMP).
- Elevated cAMP levels activate protein kinase A (PKA) and exchange protein directly activated by cAMP (EPAC), initiating downstream transcription and secretory pathways.
For a detailed breakdown of the receptor’s molecular structure and pharmacological classification, researchers can consult the GLP-1 receptor | Glucagon receptor family | IUPHAR/BPS Guide to PHARMACOLOGY.
Receptor Internalization and Endosomal Trafficking
Once activated, the GLP-1 receptor undergoes rapid internalization. This process is highly regulated and serves as a critical mechanism to control the duration and intensity of cellular signaling.
The standard pathway of internalization involves clathrin-mediated endocytosis. Upon ligand binding, G-protein coupled receptor kinases (GRKs) phosphorylate the intracellular tail of the receptor, recruiting beta-arrestin. Beta-arrestin acts as an adapter protein, facilitating the assembly of clathrin-coated pits that pinch off from the plasma membrane to form early endosomes.
In recent years, researchers have developed elegant tools to track this process in real-time. By utilizing SNAP-tag knock-in mouse models and cell-impermeable fluorescent dyes (such as LUXendin probes), scientists can selectively label surface-exposed receptors and watch them internalize. This research has revealed that different agonists promote distinct rates of endosomal trafficking:
- Exendin-4 is a highly potent inducer of internalization, rapidly recruiting up to 70–80% of surface receptors into intracellular endosomes within 30 minutes.
- Semaglutide and the dual-agonist tirzepatide exhibit more moderate internalization profiles, recruiting approximately 50% and 44% of surface receptors, respectively.
This difference in trafficking is not just a biological curiosity; it dictates whether a receptor is rapidly degraded or recycled back to the cell surface to signal again. Understanding how these cellular dynamics impact overall systemic metabolism is a key focus of ongoing research, particularly when examining how different compounds affect body composition, as discussed in The GLP-1 Guide to Not Melting Away Your Muscle.
GLP-1R Expression in Oncology: Normal vs. Tumorous Tissues
The distinct expression profile of the GLP-1 receptor in healthy tissues compared to neoplastic (tumorous) tissues has opened up exciting avenues in nuclear medicine and oncology research.
Large-scale screens analyzing hundreds of human tumors have shown that while GLP-1 receptors are virtually absent in common epithelial cancers (such as carcinomas of the breast, colon, or lung) and lymphomas, they are highly overexpressed in specific endocrine, neural, and embryonic tumors.
The tumor types showing the most robust GLP-1R expression include:
- Pheochromocytomas and Paragangliomas: These tumors show by far the highest incidence and density of GLP-1 receptors among all neoplasms analyzed.
- Insulinomas: Benign pancreatic insulin-secreting tumors express extremely high levels of GLP-1R, far exceeding the density found in normal islet cells.
- Medullary Thyroid Carcinomas (MTC): A subset of these neuroendocrine thyroid tumors express detectable levels of the receptor.
- Brain and Embryonic Tumors: Certain meningiomas, astrocytomas, and medulloblastomas also show significant receptor density.
Because normal tissues surrounding these tumors (such as the exocrine pancreas, liver, and lymph nodes) have very low or absent GLP-1R expression, there is an incredibly high tumor-to-background ratio. This contrast makes the GLP-1 receptor an exceptionally promising target for in vivo scintigraphy (using radiolabeled GLP-1 analogs like exendin-4) and targeted radiotherapy.
Researchers can read the foundational clinical study on this diagnostic potential in GLP-1 receptor expression in human tumors and human … – PubMed.
Comparing GLP-1 Receptor Expression Tissues in Humans vs. Rodents
One of the most critical hurdles in translating peptide research from bench to bedside is navigating the physiological differences between animal models and humans. While rodents are invaluable for early-stage evaluation, their GLP-1 receptor expression tissues exhibit several stark differences when compared to primates and humans.
| Tissue / Organ | Rodent Expression Profile | Human / Primate Expression Profile | Translational Significance |
|---|---|---|---|
| Thyroid C-Cells | High expression; activation drives calcitonin release and C-cell hyperplasia | Extremely low to absent; virtually no receptor detection on healthy C-cells | Explains why rodent thyroid tumor risks do not readily translate to human clinical data |
| Lung Parenchyma | Very high expression in alveolar septa | Restricted entirely to smooth muscle cells of pulmonary arteries | Rodents show direct pulmonary physiological responses that are absent in humans |
| Liver | Absent | Absent | Consistent across species; direct hepatic effects do not occur in either model |
| Visceral Arteries | Widespread expression across many small vessels | Restricted to specific renal and pulmonary arterial beds | Systemic blood pressure and vascular resistance dynamics differ between species |
These species-specific differences were elegantly demonstrated in a landmark study using a highly validated, primate-specific monoclonal antibody, which can be reviewed at (Open Access) GLP-1 Receptor Localization in Monkey and Human Tissue: Novel Distribution Revealed With Extensively Validated Monoclonal Antibody (2014) | Charles Pyke | 709 Citations. Recognizing these differences prevents researchers from making incorrect assumptions about human safety and efficacy based solely on rodent tissue data.
Frequently Asked Questions
Which human organs show the highest levels of GLP-1R expression?
In healthy humans, the highest concentrations of GLP-1 receptors are found in the pancreatic beta cells, the neurohypophysis (posterior pituitary gland) in the brain, the Brunner’s glands of the duodenum, and the supraoptic nucleus of the hypothalamus. Among tumor tissues, pheochromocytomas and benign insulinomas express the highest density of GLP-1 receptors.
Is the GLP-1 receptor expressed in the liver or skeletal muscle?
No. Rigorous transcriptomic and immunohistochemical analyses have confirmed that the GLP-1 receptor is completely absent in healthy human liver tissue, skeletal muscle, and adipose tissue. Any metabolic improvements in hepatic glucose output or muscle insulin sensitivity observed during research are indirect, downstream effects of improved pancreatic function, reduced systemic inflammation, and central nervous system signaling.
How reliable are antibodies for detecting GLP-1R in tissue samples?
Historically, GLP-1R antibodies were notorious for poor specificity, often binding non-specifically to other proteins and leading to false-positive reports of receptor presence in tissues like the liver or pancreatic ducts. Today, researchers rely on extensively validated monoclonal antibodies (such as the GLP1R-DIR antibody series), RNAscope in situ hybridization to detect mRNA directly, and transgenic reporter models (such as SNAP-tag or Cre-Lox systems) to ensure highly accurate, reproducible localization data.
Conclusion
Mapping GLP-1 receptor expression tissues reveals a highly sophisticated, tissue-enhanced network that coordinates metabolic, cardiovascular, and neurological responses. From the precise insulinotropic machinery of pancreatic beta cells to the intricate satiety pathways of the human nodose ganglion and hypothalamus, the localization of the GLP-1 receptor dictates its multi-systemic influence.
For researchers, understanding these precise cellular and subcellular boundaries is essential for designing the next generation of selective agonists, dual-agonists, and targeted radiopharmaceuticals. As we move further into the future of metabolic medicine, matching receptor distribution with functional outcomes remains one of the most exciting frontiers in peptide science.
To expand your understanding of these compounds and keep up with the latest scientific updates, explore our GLP-1 Peptides Complete Guide 2026 or browse our extensive library of resources in the GLP-1 Category.

